HIV & AIDS: mytisk virus og pseudodiagnose

Eleni Papadopulos-Eleopulos
Kap. 5:
Refleksjoner om patogenese
Sist oppdatert: 9. august 2012.
a) HIV=AIDS-skolens patogenesemodell
b) Seks sykdomsspektre betyr seks patogeneser
c) Teorien om oksidativt stress
d) Luc Montagnier om oksidativt stress og AIDS
a) HIV=AIDS-skolens patogenesemodell
”Patogenese” betyr hvordan en sykdom utvikles; dvs. ĺrsak-virkningskjeden fra primćrĺrsak til full klinisk manifestasjon. HIV=AIDS-skolen har i utviklingen av deres patogenesemodell bevart primćrfokusen pĺ immunforsvarets CD4-celler, som skal vćre den primćre mĺlcellen for det angivelige HIV-viruset (Wiki: T helper cell; Pathophysiology of HIV/AIDS).
a1: Viral Load-patogenesemodellen (1995- i dag): HIV dreper ikke vertscellen, men utlřser en (undřvendig?) hyperaktivering av immunforsvaret som etter 9-10 ĺr har brutt ned seg selv.
Pĺ begynnelsen av 1990-tallet hadde HIV=AIDS-skolen et stort forklaringsproblem nĺr det gjaldt patogenesen ved AIDS. Problemet var ĺ forklare hva som gjřr infeksjon av HIV sĺ farlig, da:
- konsentrasjonene av oppdaget HIV i sirkulerende blod og i infiserte immunceller (CD4-celler) er ualminnelig lav. Ved infeksjon av andre virus er det vanlig ĺ finne det infiserte virus i sirkulerende blod i mengder fra hundretusener til over én million pr. mL. Men for HIV fant man bare 10 pr. mL. For alle andre virus som antas ĺ forĺrsake sykdom, bryter sykdommen fřrst ut nĺr viruset har infisert ca. 1/3 av mĺlcellene. Ved virusinfeksjon er det vanlig at opp til 2/3 av alle mĺlcellene blir infiserert. Ved HIV-infeksjon fant man at bare 1:500 av CD4-cellene var infisert.
- HIV, i motsetning til andre virus, er ikke cytotoksisk (i hvert fall ikke in vitro), dvs. at den dreper ikke vertscellen.
Paul Philpott og Christine Johnson oppsummerer denne situasjonen i deres interessante artikkel Viral Load of Crap (1996): “Sĺ gitt virologiens gjeldende standarder, skulle HIV ha blitt avvist som et ikke-patogen.” HIV som patogen var altsĺ et stort mysterium bĺde nĺr det gjaldt hvor uhyre liten prosentandel av mĺlcellene som ble infisert, og det faktum at HIV ikke en gang skadet eller drepte vertscellen. HIV=AIDS-skolen trengte sĺrt en modifikasjon av patogenesemodellen. De trengte enda et krafttiltak fra pseudovitenskapen.
I 1993 publiserte Nature to interrelaterte artikler der forfatterne [Haase/Fauci] hevdet ĺ ha funnet massive mengder av HIV skjult i lymfeknutenes CD4-celler. Forfatterne hadde brukt en ny PCR-variant, kalt QC-PCR (”quantitative competetive PCR”), for ĺ telle virusene pĺ en ny mĺte. I 1995 publiserte Nature to nye interrelaterte artikler der forfatterne [David Ho og Xiping Wei] hevdet ĺ ha funnet massive mengder av HIV i det perifere blodet til AIDS-pasienter. Xiping Wei hadde i likhet med Haase/Fauci brukt QC-PCR, mens David Ho hadde brukt en ny PCR-variant kalt bDNA (”branched DNA”). Ho og Wei kombinerte deres egne pĺstander med dem til Haase og Fauci, og utarbeidet den nye patogenesemodellen for AIDS, kjent som ”Viral Load-hypotesen”. Philpott og Johnson (1996) oppsummerer denne kreative men bisarre hypotesen i syv punkter.
I Viral Load-patogenesemodellen antas HIV ĺ vćre kontinuerlig hyperaktiv i ĺ infisere nye vertsceller og mangfoldiggjřre seg selv, i hele perioden fra HIV-infeksjon til klinisk AIDS. Man formoder at HIV kontinuerlig infiserer nye CD4-celler, men at immunforsvarets CD8-celler (hvis oppgave er ĺ drepe infiserte celler) greier ĺ drepe de infiserte CD4-cellene i samme tempo som HIV infiserer dem. I denne modellen utfřrer altsĺ immunforsvarets CD4-celler et fantastisk arbeid i ĺ mangfoldiggjřre seg selv like hurtig som de blir infisert av HIV-virus og deretter drept av CD8-cellene. Og CD8-cellene utfřrer et fantastisk arbeid i ĺ drepe CD4-cellene like hurtig som de blir HIV-infisert. Det oppstĺr sĺledes en balanse i krigen der HIVs infeksjonstempo og CD8-cellenes drapstempo er likt. Denne kampen varer i gjennomsnitt i 9-10 ĺr, kjent som latensperioden. Latensperioden avsluttes nĺr CD4-cellene har blir sĺ utslitt av sin kontinuerlige mangfoldiggjřrelse at tempoet heri senkes. Resultatet er et fallende CD4-tall, som HIV=AIDS-skolen tolker som varselsignalet for at opportunistiske infeksjoner (AIDS) snart bryter ut.
Med ”Viral Load” menes den mĺlte mengden av angivelig HIV-RNA i blodet. Dette mĺ ikke forveksles med ”Viral burden”; som er den mĺlte mengden av angivelig HIV cDNA i infiserte celler.
Ifřlge Ho og Wei skal de nye PCR-variantene kunne oppdage og kvantifisere mengden av HIV-RNA i blodet pĺ et gitt tidspunkt etter HIV-infeksjon, hvilket sĺ blir basis for ĺ gi en prognose for nĺr AIDS sannsynligvis vil manifesteres. Philpott og Johnson, i artikkelen Viral Load of Crap (1996), forklarer i seks punkter hvordan Viral Load-patogenesemodellen svikter bĺde nĺr det gjelder logikk og evidens.
En kritisk gjennomlesning av de presenterte fakta i de to parene av interrelaterte artikler [Haase/Fauci 1993; Ho/Wei 1995] viser at massive mengder av angivelig HIV slettes ikke hadde blitt funnet, verken i lymfeknutenes CD4-celler eller i blodet. Forfatternes tolkninger hadde vćrt sćrdeles subjektive, og konklusjonene deres var uten substans. Forfatterne syntes ĺ vćre under den villfarelse at hvert RNA-fragment som ble funnet gjennom PCR kunne telles som en hel RNA-trĺd, og dermed at man kunne dele det totale antall av funne RNA-fragmenter med to for ĺ bestemme antall HIV-virus (gitt at HIV bestĺr av to RNA-trĺder)! PCR finner verken HIV eller HIV-genomer. PCR er ikke designet for ĺ finne eller kvantifisere HIV. PCR pĺ sitt aller beste kan ikke mer enn ĺ kvantifisere RNA-fragmenter som HIV-forskerne assosierer med HIV. Philpott og Johnson (1996), som her forutsetter at HIV er et reelt virus, skriver:
“Resultatet: De 100.000 HIV som hadde blitt talt opp ved ĺ bruke deres nye PCR-teknikk, korresponderte til mindre enn ti reelle HIV! Med andre ord: Ho og Weis forsřksgruppe hadde hatt det samme lave HIV-tallet som alltid hadde blitt observert i AIDS-pasienter! Men hvis det bare var ti HIV, hvordan kunne Ho og Wei mĺle 99.990 ekstra HIV? Noen av disse ekstra var HIV som hadde blitt nřytralisert av antistoffer; noen var defekte HIV (de som ikke hadde utviklet seg riktig); og noe var frittflytende HIV RNA. Selv om ingen av disse entitetene hadde patologisk kapasitet, skaper Viral Load-teknikken forvirring ved ĺ ta dem for ĺ vćre hele, infeksiřse virus, som er den eneste sorten som har biologisk betydning. Det meste av Ho og Weis ”Viral Load” er trolig bare en luftspeiling, et enormt antall HIV RNA-biter generert av PCR, ikke hele RNA generert av HIV. Denne forklaringen forener alle fakta: Et langsomt-repliserende virus som bare infiserer en svćrt liten prosentandel av celler (selv i lymfeknutene), og som er tilstede i infeksiřs form bare i spede konsentrasjoner…
Fauci, Haase, Ho og Wei mislyktes alle i ĺ demonstrere en rolle for HIV i AIDS fordi de mislyktes i ĺ vise at AIDS-pasienter har en betydelig mengde av HIV eller HIV-infiserte celler, eller hvorfor HIV skulle betraktes anderledes enn de mange andre virus som infiserer immunceller uten ĺ forĺrsake AIDS.”
Viral Load-patogenesemodellen er fortsatt bare en hypotese, dog den dominerende hypotesen siden 1995. Gitt at HIV nĺ har vćrt verdens mest studerte virus i 28 ĺr, er det absurd og bisarrt hvor lite HIV-forskerne ennĺ vet om dette viruset. Fřlgende tre spřrsmĺl avslřrer denne fundamentale kunnskapsmangelen:
a2: Hvorfor skulle HIV-infeksjon av max. 0,2 % av kroppens CD4-cellene utlřse krise i immunforsvaret?
Mennesket antas normalt ĺ ha 250 milliarder CD4-celler. Ifřlge HIV=AIDS-skolen blir 50-500 millioner av disse infisert ved en HIV-infeksjon, altsĺ bare 0,02 % til 0,2 % [Hazenberg et al., 2000]. Hvorfor skulle denne absolutt minimale infeksjonsgraden provosere immunforsvaret til ĺ bli hyperaktivt i 9-10 ĺr fřr det selvdestruktivt utmatter seg selv og gĺr til grunne?
a3) Hvorfor vet man ennĺ ikke med sikkerhet hvorvidt HIV dreper vertscellen eller ikke?
I Rethinking AIDS sitt dokument Rethinking AIDS Responds to the 56 ‘Errors’ (2008) kan man under de to punktene nedenfor lese en rekke nyere referanser der HIV-forskerne uttrykker sin usikkerhet om hvorvidt HIV dreper vertscellen eller ikke. De fleste mener, i motsetning til Gallo, at HIV ikke dreper vertscellen.
Nĺr man ikke en gang vet hvilken virkning det angivelige HIV-viruset har pĺ vertscellen, hvordan kan man da vite at viruset overhodet har virkning? Hvordan kan man da vite hva som er de spesifike ĺrsakene til at CD4-tallet noen ganger faller, individuelt og fra gang til gang?
a4) Hvorfor tillegger HIV=AIDS-skolen CD4-tallet sĺ stor betydning, gitt hva den biomedisinske forskningen generelt vet om CD4-tallets store individuelle og timelige variasjon og vilkĺrlighet bĺde blant friske og syke?
Nĺr CD4-tallet har falt fra normalspekteret pĺ 500-1200 celler/mL til under 200, er dette sammen med HIV-diagnosen eller én av et tyvetalls AIDS-sykdommene nok i USA til ĺ gi voksne og ungdommer AIDS-diagnosen (Wiki: CDC Classification System for HIV Infection in Adults and Adolescents). Matt Irwin viser i sin grundige artikkel fra 2001 at lavt CD4-tall ikke betyr noe som helst spesifikt, Low CD4+ T-lymphocyte counts: a variety of causes and their implications to a multifactorial model of AIDS. I mange tilfeller har ogsĺ fullstendig friske mennesker lavt CD4-tall, hvilket betyr at lavt CD4-tall ikke en gang kan fungere som varsellampe pĺ at noe er galt. Her kommer en oversettelse av artikkelens sammendrag:
“Lavt CD4-tall er assosiert med et mangfold av tilstander, inkludert mange virale infeksjoner, bakterielle infeksjoner, parasittiske infeksjoner, sepsis, tuberkolose, coccidioidomycosis, brannsĺr, intravenřse injeksjoner med fremmedproteiner, feilernćring, overtrening, graviditet, bruk av coricosteroider, normal daglig variasjon, psykologisk stress, og sosial isolasjon. Mange mennesker er fullstendig friske og har likevel et lavt CD4-tall for ingen ĺpenbar grunn.
Denne artikkelen gir et kort sammendrag av flere studier som dokumenterer lavt CD4-tall i personer med disse tilstandene. Lavt CD4-tall er sĺ vanlig at det undergraver den mest grunnleggende pĺstanden om HIV, som er at HIV forĺrsaker lavt CD4-tall. Det store flertall av personer som har blitt diagnostisert HIV-positiv har opplevd minst en av de andre ĺrsakene til lavt CD4-tall; hvilket gjřr det umulig ĺ vite om HIV er ĺrsaken.
Lavt CD4-tall forĺrsaket av noen av disse tilstandene faller ofte under 200 pr. mm3, hvilket er terskelnivĺet for ĺ kunne diagnostisere AIDS hos personer som tidligere har vćrt HIV-positive. Som eksempel, ca. 30 % av dem som har akutt lungebetennelse har vist seg ĺ ha CD4-tallet under 200. Nĺr en person som er diagnostisert HIV-positiv fĺr CD4-tallet under 200, betraktes dette som et tegn pĺ ekstrem immunsuppresjon og at HIV er i gang med ĺ řdelegge deres immunsystem. Deres lave CD4-tall vil sĺ normalt innlede et behandlingsregime der de gis flere typer antiinfektiva for ĺ forhindre infeksjoner. De vil ogsĺ bli rĺdet til ĺ begynne med proteasehemmere og DNA-kjedeterminatorer.
Fordi mange av de tilstander som forĺrsaker lavt CD4-tall er vanlig ĺ finne i personer som er HIV-positive, vil vi i denne artikkelen anbefale forsiktighet ang. behandling og diagnostiske bestemmelser pĺ basis av CD4-tallet. Behovet for ĺ utvise forsiktighet forsterkes ytterligere da noen av tilstandene, som psykologisk stress, forverres betydelig nĺr folk blir fortalt at deres CD4-tall er lavt. Dette kan resultere i at deres CD4-tall faller enda lavere. Psykologisk stress og sosial isolasjon skapes ogsĺ av ĺ fĺ HIV-positiv diagnosen og AIDS-diagnosen, hvilket kan ha kroniske virkninger pĺ CD4-tallet.
Til slutt, det almenaksepterte argumentet at HIV spesifikt angriper CD4+ T-lymfocytter vil vi ogsĺ sette spřrsmĺlstegn ved, da det virker som om lavt CD4-tall er en vanlig reaksjon pĺ mange typer av fysiologisk og psykologisk stress. Andre modifikasjoner i immunsystemets parametere som antas ĺ vćre spesifikke for HIV, slik som det omvendte CD4/CD8-forholdet, blir ogsĺ replikert svćrt bra i de tilstander som forĺrsaker lavt CD4-tall, hvilket gjřr det umulig ĺ skille ut de virkninger som tillegges HIV fra identiske virkninger forĺrsaket av andre faktorer. Nĺr flere av disse tilstander kombineres i en og samme person, som ofte er tilfelle med personer som har fĺtt HIV-positiv diagnosen, kan ekstremt lavt CD4-tall vćre et naturlig resultat.”
Matt Irwins oppfřlgerartikkel fra 2002, AIDS and the voodoo hex, gĺr nćrmere inn pĺ nocebo-effekten, dvs. hvordan psyken negativt kan pĺvirke kroppen og CD4-tallet ved ĺ fĺ HIV-diagnosen.
b) Seks sykdomsspektre betyr seks patogeneser
Kapittel 4 i dette essayet har tittelen: ”AIDS”: seks ĺrsakskomplekser, risikogrupper og sykdomsspektre. Konsekvensen herav mĺ vćre at det ogsĺ er seks spesifikke patogeneser, som for hver av risikogruppene resulterer i ervervet immunsvikt, spesifikke opportunistiske infeksjonssykdommer, og spesifikke sykdommer som ikke er relatert til immunsvikt. Vi kan anta at pĺ molekylćrbiologisk nivĺ er det snakk om forskjellige typer av ervervet immunsvikt.
Roberto A. Giraldos foreslĺtte definisjon av AIDS, i notatet The Causes of AIDS (juni 2000), gjentas her for ĺ belyse bĺde den generelle patogenese ved AIDS-spektrene og for ĺ forstĺ hvor lite som skal til pĺ det molekylćrbiologiske nivĺ fřr noe forskjellige patogeneser oppstĺr i immunforsvaret.
”Jeg foreslĺr ĺ definere AIDS som et toksisk og ernćringsmessig syndrom, det mest alvorlige og dyptgĺende av alle typer for ervervet immunsvikt, som blir forĺrsaket av mangfoldige, gjentagne og kroniske eksponeringer for immunologisk stressende agens. De stressende agens forĺrsaker degenerative immunotoksiske og immunogene effekter pĺ immuncellene og reaksjoner, med tilvekst av frie radikaler, sćrlig oksiderende agens, i alle kroppens systemer, men sćrlig i organene og vevene til immunsystemet. Disse progredierende og kontinuerlige angrep pĺ immunsystemet frembringer en kollaps i individets immunologiske funksjoner, med etterfřlgende og samtidig frembrudd av infeksjoner, svulstvekst og endret stoffskifte. Samtidig med angrepene pĺ immunsystemet, bidrar de stressende agens til ĺ endre funksjonsevnen til nćrmest talt alle andre systemer og organer. Fortsettelsen av denne prosessen kan eventuelt forĺrsake personens dřd.
De forskjellige kliniske utslag av AIDS er de direkte konsekvenser av syndromets fysiologiske og molekylćre patogenese. Fordelingen av immunologisk stressende agens varierer med risikogruppene for AIDS. Dette faktum er den primćre forklaring pĺ den store variasjon i kliniske utslag av AIDS blant dens forskjellige risikogrupper. Samtidig med at immunsystemet blir transformert, sřrger eksponeringen for stressende agens ogsĺ til skader pĺ praktisk talt alle systemer, organer og vev i kroppen. Dette er en annen faktor i forklaringen av den store variasjon i de kliniske manifestasjoner av AIDS innenfor dens risikogrupper. Ved AIDS kollapser ikke bare immunsystemet, men ogsĺ alle de andre systemene. Kort sagt, AIDS er ikke en infeksiřs sykdom, og er heller ikke seksuelt overfřrbar. AIDS er et toksisk og ernćringsmessig syndrom forĺrsaket av den alarmerende řkning av immunologisk stressende agens verden over.”
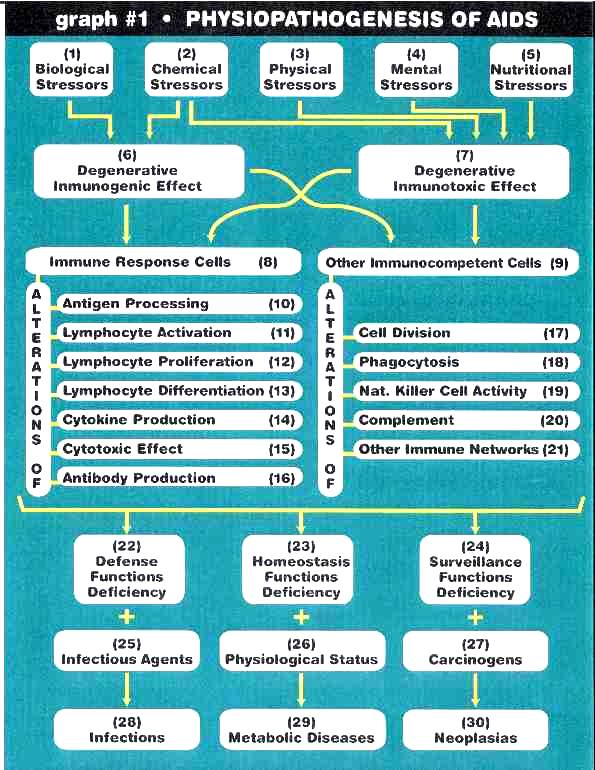
Roberto A. Giraldos patogenese-modell i notatet The Causes of AIDS (juni 2000).
Det faller utenfor dette essayets rammer ĺ diskutere mulige molekylćrbiologiske variasjoner av den generelle patogenesen som Giraldo har skissert ovenfor. I stedet gis her en generell og enkel introduksjon til oksidativt stress.
c) Teorien om oksidativt stress
Eleni Papadopulos-Eleopulos lanserte sĺ tidlig som i 1982 teorien om oksidativt stress som forklaring pĺ patogenesen ved AIDS, i artikkelen A Mitotic Theory (Journal of Theoretical Biology). Hennes videre lřp derfra som HIV/AIDS-dissident forteller hun om i artikkelen Looking back on the Oxidative Stress theory of AIDS (Continuum, 1999). Hennes teori om oksidativt stress ble opprinnelig utviklet som forklaring pĺ patogenesen ved kreft og hjerte-kar-sykdommer. Da AIDS dukket opp i media i 1981 sĺ hun at ogsĺ AIDS kunne ha oksidativt stress som patogenese, en oppfatning som hun og hennes Perth-gruppe siden har stĺtt fast ved.
Norges fremste fagekspert pĺ oksidativt stress og antioksidanter er professor i ernćringsvitenskap Rune Blomhoff ved Universitetet i Oslo (Blomhoffs webside). Han skriver at i dag publiseres over ti vitenskapelige artikler om oksidativt stress pr. dag!
La oss begynne med cellenes energiomsetning, fra lćreboken Menneskekroppen: fysiologi og anatomi (1998, s. 460):
”Cellene omsetter hele tiden energi for ĺ opprettholde sine strukturer. I tillegg trenger cellene energi for ĺ vokse, dele seg, transportere stoffer og forandre form. Cellene dřr uniddelbart hvis energiomsetningen blokkeres. Cellene er avhengige av energitilfřrsel utenfra. Grřnne planter kan utnytte energien i sollyset til ĺ bygge opp karbohydrater og vann (fotosyntesen). Dyrene fĺr tilfřrt kjemisk energi ved ĺ spise planter eller dyr. Cellene tapper de organiske molekylene i nćringsstoffene for kjemisk energi ved ĺ bryte molekylene ned i en serie kjemiske reaksjoner…”
Tappingen av energi gjennom nedbrytning av organiske molekyler skjer gjennom en serie redoks-reaksjoner (oksidasjon og reduksjon) der elektroner overfřres til stadig mer elektronegative forbindelser. Nĺr elektroner overfřres fra et atom/molekyl til et annet som er mer elektronegativt, frigjřres energi. Atomet/molekylet som avgir elektroner bli oksidert, mens atomet/molekylet som tilfřres elektroner blir redusert. Oksygenatomet er langt mer elektronegativt enn karbon- og hydrogenatomene; ved tapping av energi skjer det derfor en elektronoverfřring fra disse atomene til oksygen.
Frie radikaler er kjemisk ustabile oksygenmolekyler som bidrar til ĺ skade og svekke cellen gjennom deres mange reaksjoner. Antioksidanter nřytraliserer de frie radikaler og er vĺrt forsvarssystem mot dem. Vi fĺr vĺre antioksidanter hovedsakelig fra frukt og grřnnsaker. Her kommer et sitat fra Rune Blomhoffs artikkel Antioksidanter og oksidativt stress (2004, Tidsskriftet for den Norske Lćgeforening):
”Frie radikaler og andre reaktive oksygen- og nitrogenforbindelser dannes som et resultat av normal oksidativ metabolisme. Mange sykdommer, rřyking, miljřgifter, medikamenter, alkohol og strĺling fremmer ogsĺ danning av disse stoffene. Hvis slike reaktive molekyler ikke elimineres eller nřytraliseres av antioksidanter, kan viktige strukturelle eller funksjonelle komponenter skades, slik som lipidmembraner, lipoproteiner, signalmekanismer og RNA eller DNA (1–3). Hvis antioksidantforsvaret ikke er adekvat, vil oksidative skader akkumuleres, og tilstanden «oksidativt stress» inntreffer. En rekke studier demonstrerer at oksidativ skade og oksidativt stress er nćr knyttet til patogenese av mange tilsynelatende ikke-relaterte sykdommer (f.eks. alle inflammatoriske tilstander, iskemiske sykdommer, kreft, hemokromatose, AIDS, emfysem, magesĺr, hypertensjon, preeklampsi og nevrodegenerative sykdommer) (1–3).”
”Alle aerobe planter og dyr har etablert et omfattende forsvar mot oksidativ skade. Dette forsvaret har enzymatiske og ikke-enzymatiske komponenter som kan forebygge danning av frie radikaler, som kan nřytralisere eller fjerne frie radikaler, reparere oksidativ skade eller eliminere molekyler som har oksidativ skade (2–4). Forbindelser i kosten kan styrke antioksidantforsvaret – enten ved ĺ bidra med stoffer som nřytraliserer reaktive molekyler direkte (f.eks. vitamin C, vitamin E og betakaroten) eller ved ĺ bidra med molekyler som stimulerer eller pĺ annen mĺte bidrar til ĺ styrke cellens eget antioksidantforsvar (2–4).”
Oksidativt stress oppstĺr nĺr cellens normale funksjon trues pga. overveldende oksidativ skade. Rune Blomhoff har ogsĺ en artikkel pĺ engelsk som dekker det samme innholdet som over, men som gĺr dypere inn i biokjemien: Dietary antioxidants and cardiovascular disease (2005). Her nevner han bl.a. at studier pĺ mennesker dessverre ikke kan bekrefte at vitamin/antioksidant-supplementer har noen positiv virkning.
d) Luc Montagnier om oksidativt stress og AIDS
Luc Montagnier, som i 2008 fikk Nobelprisen for oppdagelsen av HIV i 1983, har siden 1990-tallet kommet med flere kunngjřringer, muntlig sĺ vel som skriftlig, om at han betrakter oksidativt stress som en medvirkende ĺrsak til AIDS.
I redaktřrverket Handbook of Antioxidants (Oxidative Stress and Disease) (utg. 1, 1998) har Montagnier et al. skrevet kapittelet Oxidative stress in cancer, AIDS and neurogenerative diseases. Der skriver de bl.a.
"… in AIDS pathogenesis, oxidative stress is proposed as a metabolic alteration that favours disease progression by inducing both viral replication and apoptotic death.”
I en tale Montagnier holdt i 2003 for Europaparlamentet hevdet han at ĺrsaken til AIDS er lavt CD4-tall, som skyldes apoptose (at cellen begĺr selvmord), som skyldes oksidering, som i Afrika skyldes feilernćring.
I et utvidet intervju med Montagnier fra filmen House of Numbers (2009) anbefales ĺ hřre pĺ seksjonen Co-factors, Oxidative stress and Nutrition (10:15 – 22.57 min.).

Luc Montagnier, intervjuet i House of Numbers (2009).
Tilbake til: HIV/AIDS-Innholdsside // Home