Kap. 2
HIV-viruset: Fiksjon eller realitet?
Sist revidert: 28. juli 2012.
a) Hva er et virus?
b) Om RT-enzymet og viruslignende cellepartikler
c) Ĺ isolere og karakterisere retrovirus etter gullstandarden
d) Isolerte Montagnier-teamet et retrovirus i 1983?
a) Hva er et virus?
Et virus er en mikroskopisk infeksiřs agens (virksom faktor) som bare kan mangfoldiggjřre seg inne i levende celler i en organisme (Wiki: Virus). Virus kan infisere alle typer organismer, fra dyr og planter til bakterier. Virus bestĺr av litt DNA eller RNA som er innpakket i et proteinskall. Virus varierer i střrrelse fra 10-300 nanometer (1 nanometer er like langt som ti hydrogenatomer plassert etter hverandre). Et gjennomsnittlig stort virus har ca. 1/100 av střrrelsen til en gjennomsnittlig stor bakterie. De fleste virus kan bare ses gjennom elektronmikroskop, som ble introdusert i 1931.
Den hollandske botaniker og professor Martinus Willem Beijerinck (1851-1931) regnes som virologiens far, da han i 1898 gjennom filtreringseksperimenter oppdaget at tobakkmosaikksykdommen var forĺrsaket av noe som er mindre enn bakterier. Han kalte det nye ”riket” for virus (latin for gift), og den varianten han selv hadde funnet kalte han for tobakkmosaikkvirus. Den amerikanske biokjemiker Wendell M. Stanley (1904-71) var den fřrste som i 1935 oppdaget virusets struktur og funksjon. I 1946 fikk han Nobelprisen i kjemi. To ĺr senere etablerte han Virus Laboratory ved University of California. Han kom til ĺ skrive over 150 artikler og flere břker om virus, og ble anerkjent som en verdensautoritet pĺ omrĺdet. Det var hit Peter Duesberg kom i 1964 for ĺ forske pĺ svulstfremkallende virus.
Det er tallřse forskjellige typer av virus, men hver type kan bare infisere et begrenset utvalg av vertsorganismer (bakterier, plante- og dyrearter), og her kan de igjen bare infisere helt bestemte typer vertsceller. De spesifike plante- og dyrearter risikerer sĺledes bare ĺ bli infisert av et begrenset utvalg av virus.
Klassifikasjonssystemet for virus inndeler dem i orden, familie, genus og art. Da virus mangler stoffskifte blir det ofte diskutert hvorvidt de skal regnes som levende organismer. Til tross for at de ikke har stoffskifte, sĺ er de likevel infeksiřs agens, hvilket betyr at de har en slags programstyrt agenda og at kravet til deres infeksjonsevne er absolutt. To andre typer infeksiřse agens uten stoffskifte er viroider (oppdaget i 1971) og prioner (fřrst bekreftet og isolert pĺ 1980-tallet).
Gitt definisjonen av virus, betyr det at humant endogent retrovirus (HERV), som utgjřr nesten 8 % av det menneskelige genom, ikke kvalifiserer som virus (Wiki: Endogenous retrovirus). HERV er DNA-nukleotidsekvenser av tidligere retrovirus, som nĺ er en integrert del i vertscellens genom og som blir automatisk replisert nĺr celler deler seg og repliserer sitt eget DNA. HERV mangler bĺde agenda og infeksjonsevne.
Virus kan bare lage kopier av seg selv ved ĺ infisere celler. Sĺ fort viruset har kommet pĺ innsiden av cellen, utnyttes cellens eget maskineri til ĺ demontere viruset i komponenter. Komponentene blir mangfoldiggjort, og sĺ montert sammen igjen til nye virus. Noen typer virus dreper cellen for ĺ komme seg ut, andre typer virus forlater den uskadd.
b) Om RT-enzymet og viruslignende cellepartikler
I 1970 ble enzymet revers transkriptase (RT) oppdaget i kreftcellekulturer (Wiki: Reverse transcriptase). RT-enzymet transkriberer RNA til cDNA (”c” stĺr for komplementćrt), hvilket er det motsatte (revers) av det som frem til 1970 var molekylćrbiologiens sentrale dogme, at den eneste mulige transkripsjon er fra DNA til mRNA (”m” stĺr for messenger). Forskerne antok at cellene i cellekulturen hadde blitt cancerřse fordi de hadde blitt infisert av ”RNA tumor virus” og at det var fra dem RT-enzymet kom. Oppdagelsen av dette enzymet resulterte i at Howard Temin og David Baltimore i 1975 ble belřnnet med Nobelprisen. Da forskerne antok at RT-enzymet kom fra ”RNA tumor virus”, ble disse virus omdřpt til retrovirus (Wiki: Retrovirus). Oppdagelsen av RT-enzymet ga ny nćring til virusjegernes vĺte drřm om ”onkovirus” som kan forĺrsake kreft i menneskeceller.
Det er RT-enzymets funksjon i retrovirus som har gitt retroviruset dets navn, og som gjřr retrovirus til retrovirus. Kort tid etter 1970 ble det imidlertid klart at RT-aktivitet er et utbredt fenomen i den biologiske verden. I 1971 ble det kjent at RT-aktivitet er vanlig i et stort antall animalske celletyper, samt i bakterier (Mirko Beljanski).
I cellekulturer er det fullt av viruslignende partikler, RT-aktivitet, frittflytende RNA-sekvenser og DNA-sekvenser. Nĺr man har prřvd ĺ isolere infeksiřse retrovirus (som f.eks. HIV), og etter sentrifugering finner ved 1,16 bĺndet viruslignende partikler, RT-aktivitet og diverse DNA/RNA-fragmenter, sĺ er disse funnene alt for generelle til at man kan fastslĺ at et virus har blitt funnet. Isoleringsprosedyren er ennĺ langt fra fullfřrt.
c) Ĺ isolere og karakterisere retrovirus etter gullstandarden
Virologene har en konsensus-forstĺelse av hva som utgjřr den ideelle og fullstendige prosedyre for ĺ isolere og karakterisere et retrovirus. Prosedyren bestĺr av en sekvens av trinn som skal gjřres i en bestemt rekkefřlge. Dersom ett eller flere av trinnene utelates, svekkes konklusjonsgrunnlaget. Da řker faren for at man trekker ugyldige konklusjoner. Konsensus-forstĺelsen av den ideelle og fullstendige prosedyre for ĺ isolere og karakterisere retrovirus har aldri blitt nedtegnet i et eget konsensus-dokument. Perth-gruppen refererer til konsensus-forstĺelsen med begrepet ”gullstandarden”, og hevder at betydningsfulle punkter i gullstandarden kom til uttrykk ved Pasteur Institute, Paris, i tilknytning til en střrre konferanse som ble holdt i 1973. Denne konferansen dreide seg utelukkende om metodene for ĺ isolere retrovirus. Perth-gruppen refererer generelt til 1973-utgaven av Spectra, og spesielt til to dokumenter der enkelte nřdvendige trinn i isoleringsprosedyren ble poengtert (her).
For ĺ identifisere et hittil ukjent virus, mĺ viruset isoleres og karakteriseres. Verbene ”isolere” og ”rense” brukes her som synonymer, dvs. at man isolerer viruset fra alle omkringliggende partikler. La oss se nćrmere pĺ hovedtrinnene i prosedyren:
1) Fra celledyrking til sentrifugering. Virologen dyrker noen celler som antas ĺ vćre infisert med viruset. Cellene dyrkes i et flytende medium, slik at de nyskapte virusene kommer ut i mediumet nĺr de forlater cellen. Nĺr virologen antar at mediumet inneholder nok virus, er neste skritt ĺ overfřre en drĺpe fra mediumet til toppen av en sukkerlřsning inne i et sentrifugerřr. Sukkerlřsningen har pĺ forhĺnd blitt forberedt slik at den har lavest tetthet řverst i rřret og hřyest tetthet nederst i rřret. Rřret blir sĺ sentrifugert i ekstremt hřye hastigheter i flere timer. Dette fĺr partiklene i drĺpen til ĺ synke nedover i sukkerlřsningen helt til hver av dem stoppes ved det ”bĺnd” der de har samme tetthetsgrad som sukkerlřsningen. Alle retroviruspartikler konsentreres ved et helt bestemt bĺnd, nemlig 1,16 gram/milliliter. Dette bĺndet kan sĺ ekstraheres og fotograferes med et elektronmikroskop.
2) Isolere infeksiřse retroviruspartikler fra andre partikler. Selv om alle infeksiřse retroviruspartikler etter lćreboken konsentreres ved 1,16-bĺndet, sĺ er de ikke alene om ĺ havne der. Flere andre typer partikler havner ogsĺ ved dette bĺndet. Disse viruslignende cellepartiklene, inkludert cellulćrt DNA, RNA og proteiner, kan til en viss grad skjelnes fra retroviruspartikler ved ĺ titte i et elektronmikroskop.
RT-enzymet konsentreres ogsĺ ved 1,16-bĺndet. Mange retrovirologer har trodd, og tror visst ennĺ, at det er nok ĺ oppdage RT-aktivitet for ĺ kunne fastslĺ tilstedevćrelsen av et infeksiřst retrovirus. For noen retrovirolger er oppdagelsen av RT-aktivitet til og med ensbetydende med ĺ ha funnet HIV! RT-aktivitet er som nevnt verken spesifikt for HIV, retrovirus eller for virus generelt.
For ĺ vite at et infeksiřst virus har blitt funnet, mĺ partiklene som har blitt isolert fřrst overfřres til en jomfruelig cellekultur der de mĺ demonstrere at de er istand til ĺ infisere celler. Partiklene mĺ sĺ vćre istand til ĺ mangfoldiggjřre seg i cellen, for sĺ ĺ komme ut i det flytende mediumet. Partiklene i denne sekundćre kulturen skal ha eksakt samme form og egenskaper som de opprinnelige partiklene. Fřrst nĺr prosedyren over har blitt gjennomfřrt til punkt og prikke, kan man vćre sikker pĺ ĺ ha isolert et infeksiřst virus og ikke bare viruslignende cellepartikler.
3) Karakterisere retroviruset. Ĺ karakterisere et retrovirus innebćrer ĺ finne dets unike kjennetegn: Dets spesifikke RNA og proteiner, samt dets eksakte střrrelse, vekt og form. For retrovirus mĺ det ogsĺ bekreftes at et av proteinene er RT-enzymet.
4) Finne antistoffer med hřyest mulig spesifisitet. Et antistoff er et stort Y-formet protein produsert av immunforsvaret for ĺ kunne reagere med og binde seg til et spesifikt overflateprotein pĺ en inntrenger (et antigen). Inntrengeren, f.eks. en bakterie eller et virus, blir altsĺ ikke ”gjenkjent” i sin helhet, bare gjennom de overflateproteiner som tidligere har resultert i dannelsen av antistoffer. Da flere typer bakterier og virus har de samme overflateproteiner, kan det samme antistoffet altsĺ reagere pĺ flere typer inntrengere; det kryssreagerer.
Nĺr man skal undersřke om en organisme er infisert av et bestemt virus, er det som regel enklere ĺ se etter virusets antistoffer ved hjelp av en antistofftest enn ĺ se etter selve viruset. Dette forutsetter imidlertid at man kjenner antistoffenes spesifisitet, som igjen forutsetter at viruset virkelig har blitt isolert etter gullstandarden. Hvis et bestemt antistoff er 100 % spesifikt for en bestemt inntrenger, sĺ betyr det at antistoffet reagerer pĺ et bestemt overflateprotein pĺ inntrengeren som ingen andre inntrengere har.
Nĺr et virus har blitt isolert etter gullstandarden, kan man begynne ĺ identifisere dets antistoffer. I beste fall vil noen antistoffer vćre spesifikke, mens andre antistoffer vil vćre ikke-spesifikke. Dersom et virus ikke har blitt isolert etter gullstandarden, kan man ikke bestemme tilstedevćrelsen av dette ved hjelp av antistoffer, for man vet ikke hva de forskjellige antistoffene reagerer pĺ i det enkelte tilfelle. Dette prinsippet kan synes selvsagt, men Eleni Papadopulos-Eleopulos gir eksempler pĺ at verdens fremste retrovirologer – som Duesberg, Montagnier og Gallo – enten ikke har forstĺtt dette prinsippet godt nok, eller har valgt ĺ ignorere dette prinsippet nĺr det har passet dem.
* * *
Nĺr de her fire nevnte trinnene i isolering og karakterisering av et virus har blitt utfřrt, kan infiserte cellekulturer sendes til andre virologer og laboranter verden over. Disse kan da gjenta eksperimentene og bekrefte for hverandre at alt er i skjřnneste orden. Nĺr det gjelder HIV, har virologene verden over aldri hatt noe annet ĺ forholde seg til enn tvilsomme pĺstander om at 8-10 proteiner, som alle mennesker synes ĺ ha i forskjellige mengder, er komponenter av HIV-viruset. Hele HIV-AIDS-forskningen og hele HIV-AIDS-industrien frem til i dag er basert pĺ hva Montagnier pĺsto han gjorde i sitt laboratorium i 1983. HIV har aldri blitt isolert etter gullstandarden etter dette.
Det er naturlig ĺ spřrre om andre virus har blitt isolert etter gullstandarden. Jeg stilte dette spřrsmĺlet til Valendar Turner i Perth-gruppen den 16. juli 2012, og fikk fřlgende svar samme dag:
”Dear Mr Myhre,
We don't have any information on how other viruses have been isolated. We haven't studied this. But as Robin Weiss says on our website you don't prove the existence of one virus by pointing to the existence of others. So it's all about HIV.
We would be interested to see the step by step laboratory procedure the debunkers consider proves the existence of a virus. Perhaps you could put one or two on the spot.
We suggest you study read the commentary to The Emperor's New Virus? at
www.theperthgroup.com/OTHER/ENVCommentary.pdf
and then watch the video.
One has to understand what isolation is about and the difference with purification. To the man in the street the words mean the same thing. But not to virologists.
Best wishes,
Valendar Turner”
d) Isolerte Montagnier-teamet et retrovirus i 1983?
I mai-nummeret 1983 av Science ble Montagnier-teamets artikkel trykt, om isolering av et retrovirus hos en pasient med risiko for ĺ utvikle AIDS (abstract). Forfatterne uttrykte at virusets rolle som evt. ĺrsak til AIDS ”gjenstĺr ĺ bestemme”. I mai-nummeret 1984 av Science ble fire artikler av Gallo-teamet trykt samtidig, der de hevdet: 1) ĺ ha isolert et retrovirus, 2) at dette retrovirus er ĺrsaken til AIDS. To av de fire artiklene til Gallo-teamet inneholdt ordene ”isolation… of cytopathic retroviruses…” Som Perth-gruppen pĺpeker, i virologien er betydningen av ”ĺ isolere” ĺ bevise tilstedevćrelsen av et virus. Dersom et virus har blitt isolert, har virusets tilstedevćrelse blitt bevist. Det er sĺledes ingen tvil om hva Montagnier-teamet og Gallo-teamet mente og hevdet de hadde prestert.
Perth-gruppen har hele tiden hevdet at Montagnier-teamet ikke isolerte et retrovirus vĺren 1983. Dersom et retrovirus som senere fikk navnet HIV ikke ble isolert da, har retroviruset ved navn HIV heller aldri senere blitt isolert. En nćrmere analyse av Montagnier-teamets egne beskrivelser av deres isolasjonsprosedyre vĺren 1983, basert pĺ deres egne artikler samt hva de i flere intervjuer har uttalt, viser at relatert til gullstandarden var deres isolasjonsprosesdyre sćrdeles mangelfull. De trakk dermed konklusjoner som det overhodet ikke var grunnlag for ĺ trekke. Deres faktiske funn kvalifiserer ikke en gang til ĺ konkludere at de fant virus overhode. Perth-gruppen hevder at det eneste de to teamene fant var cellulćre proteiner og partikler.
Som Val Turner pĺpeker i eposten over, den beste introduksjonen til dette molekylćrbiologisk temmelig teknisk-detaljerte emnet, er fřrst ĺ se Brent Leung dokumentarfilm The Emperor’s New Virus? - An Analysis of the Evidence for the Existence of HIV (2011), og deretter sette av noen timer (eller dager) til grundig ĺ studere Perth-gruppens kommentardokument, The Emperor's New Virus? Commentary by the Perth Group 20th September 2011 (2011).
Her er noen av hovedpunktene fra Perth-gruppens kommentardokument:
- Montagnier-teamets ”isolasjon” av HIV:
I det fřrste eksperimentet ble lymfocytter fjernet fra en hoven lymfeknute i nakken til en homoseksuell mann med navnekoden BRU. 15 dager senere ble RT-aktivitet oppdaget i cellekulturen, hvilket teamet tolket som tilstedevćrelsen av et retrovirus.
I det andre eksperimentet ble lymfocytter fra en frisk blodgiver tilfřrt den fřrste cellekulturen. Pĺ ny ble RT-aktivitet oppdaget. Uten ĺ kunne vite hvem (BRU eller den friske) som cellene med RT-aktivitet kom fra, konkluderte teamet at de nĺ hadde pĺvist retrovirusets replikasjonsevne.
I det tredje eksperimentet ble noe av cellekulturen fra det andre eksperimentet tilfřrt uinfiserte lymfocytter fra navlestrengsblod fra to nyfřdte barn. Ogsĺ i dette eksperimentet ble RT-aktivitet oppdaget. Montagnier-teamets eneste publiserte EM-bilde kom fra dette eksperimentet, og var et bilde av cellekulturens overflatelag, ikke av 1,16-bĺndet! EM-bildet er ifřlge to ledende EM-eksperter, Hans Gelderblom og Reinhard Kurth, for utydelig og udetaljert til at man kan konkludere noe som helst. Montagnier-teamet mente at EM-bildet viste et retrovirus tilhřrende en annen underfamilie enn den som HIV nĺ er klassifisert under. Uansett hva EM-bildet viste; det kom nřdvendigvis ikke, og sannsynligvis ikke, fra BRU!
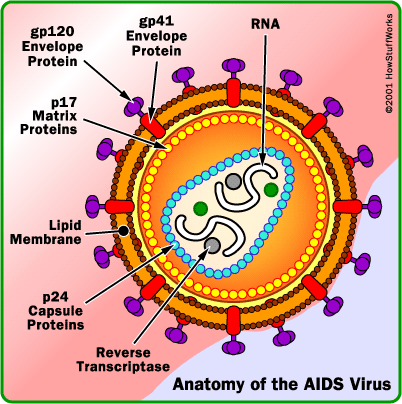
Da Montagnier-teamet konkluderte at de hadde isolert et virus, kom neste oppgave: Ĺ karakterisere viruset. Dette er absolutt nřdvendig hvis man řnsker ĺ gjřre krav pĺ ĺ ha oppdaget et nytt virus. Ca. 60 % av vekten av retrovirus bestĺr av proteiner, og ca. ti proteiner er vanligvis involvert. Vi kunne dermed forvente at Montagnier-teamet ville identifisere ca. ti virale proteiner, og noen av dem burde vćre unike for retrovirus.
Montagnier-teamet utfřrte et protein-antistoff-eksperiment i BRU-serumet, og oppdaget at tre forskjellige antistoffer reagerte med tre forskjellige proteiner. De konkluderte at p24 (som de opprinnelig oppfattet som p25) var et retroviralt protein, og at p41 (som de opprinnelig oppfattet som p45) og p80 var cellulćre proteiner. Elektroforese-bildet som ble trykt viste imidlertid at de oppdagete proteiner bare hadde blitt funnet i en celle-ekstrakt fra BRU-serumet, mens i lřsningen med det ”isolerte viruset” hadde ingen av proteinene blitt funnet!
Journalisten Djamel Tahi intervjuet Montagnier i 1997, Interview Luc Montagnier, og spurte hvorfor bare ett EM-bilde av det isolerte retroviruset hadde blitt trykt. Montagnier svarte:
”Vi sĺ noen partikler, men de hadde ikke den typiske morfologien til retrovirus. De var svćrt forskjellig.”
Da HIV ikke er klassifisert som ”atypisk retrovirus” (en slik taksonomisk betegnelse finnes ikke), er det enklest ĺ anta at hva Montagnier sĺ i sitt ”rensede virus” var ubestemmelige partikler som manglet retroviral morfologi. I intervjuet med Tahi innrřmmet Montagnier til og med: ”Jeg gjentar, vi renset ikke”.
Perth-gruppen konstaterer:
- At to av de tre proteinene som ble funnet ble avvist som cellulćre, betyr at isolasjonen var kontaminert. Hvordan kunne Montagnier-teamet da vite at p24 var viralt?
- Hvor ble de ni andre proteinene av? Det finnes ikke retrovirus med bare ett protein.
- Hvorfor greide ikke Montagnier-teamet en gang ĺ finne RT-proteinet (p66+p51), da oppdagelsen av RT-aktivitet var selve basisen for deres konklusjon om ĺ ha funnet et retrovirus? Uten RT-proteinet kan de ikke ha funnet et retrovirus! Dette betyr igjen at RT-proteinet som sto bak RT-aktiviteten i de tre eksperimentene var cellulćrt.
Til tross for den sćrdeles utilfredsstillende isolasjonsprosedyren, og de merkelige inkonsekvente slutningene nĺr det gjaldt p24, har p24 fortsatt i dag status som dét unike HIV-proteinet. Gallo-teamet konkluderte at p41 (Montagniers p45) ogsĺ er et HIV-protein.
- Problemet med ĺ bekrefte HIV-virusets infeksjonsevne og gp120:
I dag er alle ”HIV”-eksperter enige om at HIV-virusets overflateprotein gp120 er absolutt nřdvendig for at viruset skal kunne binde seg til en reseptor pĺ CD4-cellen, og sĺledes kunne infisere CD4-cellen. Formelen kan uttrykkes slik:
Ingen gp120 = ingen infeksjonsevne = ingen replikasjon = ingen ny virus.
gp120-proteinene skal se ut som pigger (pĺ engelsk kalt ”knobs” og ”spikes”). Det store problemet og mysteriet for HIV=AIDS-skolen er at ingen HIV-forskere ennĺ har kunnet bevise eksistensen av pigger pĺ cellefrie HIV-partikler. Da kan ikke disse partiklene vćre infeksiřse. Da kan de heller ikke vćre virus! Pr. idag har ingen publisert EM-bilder av cellefrie partikler med retroviral morfologi og pigger.
Kuznetsov et al. fikk i 2003 publisert artikkelen Atomic force microscopy investigation of human immunodeficiency virus (HIV) and HIV-infected lymphocytes (Journal of Virology), der bilder av angivelig HIV med den nye bildeteknologien ”atomic force microscopy” (AFM) for fřrste gang ble publisert. AFM-bildene av angivelig HIV viser at gp120, som skal danne infeksjonspiggene pĺ HIV, manglet piggformen. Forfatterne er av den oppfatning at den lille uthevningen som synes, er en bildeartefakt og ikke noe reelt ved viruset. I artikkelen til Ping Zhu et al. (2006), Distribution and three-dimensional structure of AIDS virus envelope spikes (Nature 441, 847–852), kunne ikke forfatterne bekrefte med sikkerhet at HIV har pigger.
- Elektronmikroskop-bilder:
Frem til mars 1997 hadde ingen HIV-forskere publisert et eneste EM-bilde av den materien som samlet seg ved 1,16-bĺndet. De to fřrste EM-studiene av 1,16-materien var:
I begge studiene ble det hevdet at det meste av materien, opp til 80 %, var cellulćre partikler (proteiner, retroviruslignende partikler, DNA og RNA). Spřrsmĺlet var hva som var forfatternes grunnlag for ĺ hevde at det řvrige materialet kom fra retrovirus. EM-bildene viser ikke partikler med retroviral morfologi, sĺ det er ikke en gang grunnlag for ĺ hevde at retroviruslignende partikler ble funnet.
Begge artiklene avslřrer betydelig usikkerhet i det meste nĺr det gjelder ĺ kunne skjelne HIV fra alt det cellućre ”skittet” ved 1,16-bĺndet. Forfatternes tolkninger av EM-bildene er heller ikke tillitsvekkende. Det synes ikke ĺ vćre noen begrensninger nĺr det gjelder hvilke střrrelser eller former HIV kan anta. Perth-gruppen (2011):
“In other words, the Gluschankof and Bess papers affirm that up until 1997 none of the HIV experts knew what particles, if any, their “purified viruses” contained and if present their morphologies or degree of purity. In 1997 these authors showed that “purified HIV” consists almost entirely of cellular structures (“microvesicles”) with a few other particles that lack several, key defining features of lentiviruses. Yet these data made no difference to the “overwhelming scientific consensus”. HIV experts kept using this material (proteins and RNA) as the basis of diagnosis and treatment, just as they had before. In the words of John Moore, this is “as bizarre as it gets”."
Studien til Bess et al. (1997) inkluderer et elektroforese-bilde av oppdagete proteiner i de ”HIV-infiserte” cellene. De fleste ”HIV-proteinene” mangler, og det gis ingen begrunnelse for hvorfor vi skal godta pĺstanden om at de ”HIV-proteiner” som ble funnet var virale fremfor cellulćre. Elektroforese-bildet viser ogsĺ at samtlige proteiner som var ĺ finne i de infiserte cellener, ogsĺ var ĺ finne i de uinfiserte cellene, dog i langt mindre mengder. Det var altsĺ bare en kvantitativ forskjell, ikke en kvalitativ forskjell, mellom proteinene i de infiserte og de uinfiserte celler. Hva var da grunnlaget for ĺ hevde at proteinene i de infiserte cellene var HIV-proteiner, mens de samme proteinene i de uinfiserte cellene var… ?
Perth-gruppen kontaktet Bess og medforfatterne for ĺ fĺ vite hva som var deres grunnlag for konklusjonen at tre av proteinene som ble funnet var HIV-proteiner, da de bare hadde proteinenes omtrentlige molekylćrvekt ĺ gĺ ut ifra. Svaret var at en av fagfellene som hadde vurdert manuset, hadde bedt om en slik vinkling. Bess innrřmmet at det var ingen evidens for at disse tre proteinene var HIV-proteiner.
Alexander Russell utlovet i april 2002 en dusřr pĺ 10.000 pund til den fřrste person som kunne frembringe et EM-bilde av HIV pĺ basis av syv kriterier satt opp av EM-veteranen Etienne de Harven (her). De syv kriteriene som de Harven spesifiserte, er pĺ ingen mĺte sćre eller ualminnelige. De syv kriteriene břr vćre en enkel sak ĺ oppfylle for en hver forsker, student i mikrobiologi eller lab-tekniker med tilgang til et laboratorium og en HIV-diagnostisert person som ikke gĺr pĺ anti-HIV-medisiner.
Etienne de Harven skrev i artikkelen Human Endogenous Retroviruses and AIDS Research: Confusion, Consensus, or Science? (2010):
“Alle bildene av partikler som angivelig representerer HIV, og som blir publisert i vitenskapelige sĺvel som almenne tidsskrifter, er basert pĺ EM-studier av cellekulturer. De viser aldri HIV-partikler som kommer direkte fra en HIV-pasient. Bildene er alltid forskjřnnet av computerbilde-rekonstruksjon, med attraktive farger og raffinerte 3D-effekter. Den endelřse publiseringen i media verden over av disse elegante artefaktene har bidratt mye til ĺ overbevise bĺde forskere og almenheten om eksistensen av HIV...”
- Ingen enighet om klassifikasjon av HIV-partikler:
Det rĺder stor usikkerhet om klassifikasjonen av HIV-partikler. Perth-gruppen skriver:
“Yet, over the years, including at least up until 2005, different laboratories have classified the HIV particle as type-C, type-D and lentivirus particles, that is, into two subfamilies and three genera. This is no different from reporting one and the same mammal as human, a chimpanzee and an orang-utan.”
- Fravćret av kontrollgrupper:
Det meste av HIV-forskning har et oppsiktsvekkende og skrikende fravćr av kontrollgrupper. Med det menes testing av cellekulturer fra personer som ikke har AIDS-diagnosen, men som har demografiske, kliniske og laboratoriemessige abnormaliteter lik gruppen med AIDS-pasienter. Hensikten med bruk av slike kontrollgrupper vil vćre ĺ bestemme hvorvidt RT-aktivitet, retroviruslignende partikler og antistoff/antigen-reaksjoner rapportert av Montagnier-teamet og Gallo-teamet virkelig skyldes retrovirus-infeksjon, eller kan ha andre ĺrsaker.
Kilder og ressurser:
* Brent Leung (2011): The Emperor’s New Virus? - An Analysis of the Evidence for the Existence of HIV. [Dokumentarfilm, YouTube, 1t:36min.]
Dette er Leungs oppfřlger av dokumentarfilmen House of Numbers (2009). I denne oppfřlgeren er det Perth-perspektivet (i motsetning til Duesberg-perspektivet) som frontes: At det ikke er evidens for HIV-virusets eksistens, da HIV aldri har blitt isolert pĺ en tilfredsstillende og adekvat mĺte.
* Perth Group (2011): The Emperor's New Virus? Commentary by the Perth Group 20th September 2011. Dette er Perth-gruppens kommentardokument (54 sider) til Brent Leungs dokumentarfilm.
Tilbake til: HIV/AIDS-Innholdsside // Home